 |
|
Curso de Botánica
Presentación
Anatomía vegetal
Taxonomía vegetal
Adaptaciones vegetales
Sucesiones vegetales
Vegetación de la península Ibérica
Itinerarios botánicos
Introducción
Albelda
Castillonroy
Baldellou
Congosto de Baldellou
Purroy de la Solana
Barranco de Gabasa
Peralta de la Sal
San Esteban de Litera
Azanuy
Alcolea de Cinca
TAXONOMÍA VEGETAL.
Este capítulo está dedicado a clasificar y caracterizar los distintos grupos de plantas, especialmente aquellas que no son angiospermas. También se repasan los hongos y los líquenes, pese a no ser vegetales.
Las plantas terrestres proceden de antecesores de las actuales algas verdes, las cuales consiguieron abandonar el medio acuático y colonizar el medio terrestre. La evolución siguió dos líneas, una de constitución más simple, los denominados briofitos (musgos y hepáticas), que carecen de tejidos conductores, y otra, con estructuras más complejas, los cormofitos o plantas superiores, que presentan tejidos vasculares, raíces, tallo y hojas. Estas características estructurales de las plantas superiores, les permitió adaptarse y responder más adecuadamente a las condiciones cambiantes del nuevo medio terrestre colonizado.
Los talófitos son organismos cuya estructura histológica y morfológica corresponde al talo, es decir, su cuerpo u órgano vegetativo está compuesto de una masa celular indiferenciada y no existen en ellos verdaderos órganos conductores aunque, en los casos de talos más complejos, pueden existir unas células centrales, algo más permeables y diferenciadas, para la conducción del agua. Algunos talófitos poseen una estructura que refleja, en un nivel inferior, la apariencia de cormo, como sucede en algunas algas rodofíceas y feofíceas. Presentan un órgano, el cauloide, que, a modo de tallo, sirve de sostén al vegetal. También poseen órganos de fijación o rizoides que cumplen alguna de las funciones de la raíz y órganos laminares fotosintéticos o filoides con apariencia, pero no con estructura, de hojas. Por su forma, el talo puede ser filamentoso, laminar, macizo, etc., aspecto que depende de en que dirección se produce su crecimiento.
Los talófitos reúnen seres autótrofos (como las algas) o bien heterótrofos (como
los hongos); incluyen también a los líquenes. Pueden ser unicelulares o
pluricelulares.
Los protocormofitos son plantas no vasculares (briofitos), carentes de vasos
conductores de savia, con una organización intermedia entre las talófitas y las
cormófitas. Son plantas criptógamas, es decir, carentes de flores. Se
caracterizan por poseer clorofila y por su alternancia de generaciones, en la
que las dos fases (gametófito y esporófito) permanecen íntimamente unidas.
Agrupa a las hepáticas y a los musgos.
El término cormofito se utiliza para referirse a vegetales pluricelulares altamente diferenciados. Sus células están agrupadas en tejidos. El cormo es un nivel de organización que presenta raíz, tallo y hojas, en cuyas partes está diferenciado el cuerpo vegetativo. Son los vegetales superiores, adaptados a la vida terrestre, las plantas vasculares, traqueofitos o rizofitas, que se independizaron del medio acuático hace más de cuatrocientos millones de años, derivando de sus antepasados las algas verdes. En su adaptación al medio terrestre sufrieron importantes trasformaciones estructurales, tales como el desarrollo de raíces que les permitieron fijarse al suelo, y del tallo para sustentar las hojas y los órganos reproductores, el cual está dotado además de un sistema de tejidos conductores de la savia para transportar y sintetizar los nutrientes (xilema y floema) Actualmente el grupo dominante y más perfeccionado es el de las espermatofitas.
Las espermatofitas son las plantas provistas de semillas. Poseen un elevado grado de organización, con tallo, hojas y raíz, y un sistema vascular muy desarrollado. El esporófito domina sobre el gametófito que es muy reducido. Estos vegetales, además, siempre tienen flores (fanerógamas) Comprenden dos grupos: Gimnospermas y Angiospermas.
Las Gimnospermas presentan carpelos no diferenciados en ovario, estilo y estigma, y cuyos óvulos y semillas no se forman en cavidades cerradas. Sus flores son siempre unisexuales y son plantas leñosas de porte variado, árboles o arbustos. Comprenden cuatro clases con representantes vivos en la actualidad: cicadinas, ginkgoinas, gnetinas y coníferas; y tres clases con representantes fósiles: las pteridospermas, las bennettiatales y las cordaitales.
Las Angiospermas son las vulgarmente llamadas plantas con flores, que constituían, junto a Gnetofitos y Gimnospermas, las antiguas Fanerógamas. Las Angiospermas son de la flora actual el grupo dominante y el más diversificado, en cuanto a adaptaciones y biotipos. Se diferencia de las Gimnospermas por tres características: los carpelos o escamas ovulíferas envuelven a los óvulos en su totalidad, transformándose en fruto y semillas tras la fecundación; los órganos reproductores pueden ser bisexuales agrupados en flores típicas, y presentar hojas florales estériles (periantio) dedicadas sólo a la protección de la verdadera flor; realizan una doble fecundación, con desarrollo de un tejido alimenticio que rodea el embrión llamado endosperma. Las Angiospermas comprenden plantas herbáceas anuales y perennes, arbustos y árboles, trepadoras, epífitas, parásitas y saprofitas, acuáticas y terrestres. Presentan hojas bien desarrolladas, con función fotosintética, adaptadas en forma y estructura a las condiciones del hábitat. Se las divide en dos grandes grupos con características de clase: dicotiledóneas y monocotiledóneas.
Las dicotiledóneas presentan semillas con dos hojas embrionarias (cotiledones). Sus tallos pueden crecer en espesor gracias a la actividad del cambium. La raíz es pivotante (eje principal más engrosado y con larga vida). Las flores, según las familias, son pentámeras o tetrámeras. Teniendo en cuenta la estructura del periantio se las clasifica en: 1) monoclamídeas o apétalas, 2) dialipétalas y 3) simpétalas o gamopétalas. Las hojas suelen ser pecioladas y tienen la nerviación de tipo reticular. Por lo que respecta a su organización interna presentan los vasos conductores abiertos y dispuestos en círculo, lo que las diferencia de las monocotiledóneas. Las dicotiledóneas incluyen a la mayoría de las plantas superiores actuales aunque las monocotiledóneas son más evolucionadas.
Por su parte, las plantas monocotiledóneas poseen embriones con un único cotiledón. Su raíz principal es de corta duración, haces conductores dispersos por la sección transversal del tallo, carentes de cámbium funcional (no hay crecimiento secundario en grosor) y predominio de las especies herbáceas poco ramificadas.
Los briofitos, a diferencia de los hongos, son plantas verdes autótrofas. Se
trata de los primeros vegetales "puente" entre la vida acuática y terrestre. Se
clasifican en dos grandes clases: Hepáticas y Musgos. Se trata de pequeñas
plantas adaptadas a hábitats húmedos; aunque pueden presentar estructura talosa, se encuentran en general
diferenciadas externamente en tallitos y hojitas, fijándose al suelo mediante
estructuras rizoides que en ocasiones son capaces de absorber agua.
húmedos; aunque pueden presentar estructura talosa, se encuentran en general
diferenciadas externamente en tallitos y hojitas, fijándose al suelo mediante
estructuras rizoides que en ocasiones son capaces de absorber agua.
Los periodos de desecación los mantienen vegetativos, sea cubriendo superficies rocosas, troncos, paredes, tejados, etc., reviviendo después cuando las condiciones de humedad les favorece. Dado que los briofitos almacenan agua entre sus hojas, otros vegetales pueden aprovecharla; por este motivo suelen ser los primeros organismos que se instalan sobre el sustrato y superficies rocosas, aportando de esta forma materia orgánica para contribuir a mantener las siguientes sucesiones vegetales.
Se pueden reproducir sexual o asexualmente. La sexual se realiza mediante la unión de dos gametos (ovocélula y espermatozoide), siempre en un medio húmedo, en el interior del arquegonio (gametangio femenino). La reproducción asexual se realiza mediante fragmentación del gametofito, o por gemación del protonema. También se pueden formar nuevos individuos a partir de los llamados propagulos, consistentes en unos ramitos especialidados que pueden existir en los musgos. Los briofitos presentan una clara alternancia de generaciones, con un esporofito diploide y un gametofito verde y haploide (el protonema)
El protonema del musgo, consistente en filamentos verdes creciendo muy ramificados en sentido vertical, y conteniendo gran cantidad de cloroplastos, representa ese tapiz verdoso que podemos observar sobre las rocas o lugares húmedos. El gametofito de los musgos, que es siempre foliáceo, y en posición erecta o tumbada, puede llegar a alcanzar el medio metro de longitud.
Por su parte, las hepáticas, presentan un aparato vegetativo aplanado (gametofito), en aquellas que son rastreras como las hepáticas talosas; o folioso, con hojitas dispuestas dorsiventralmente, como en las hepáticas foliosas. Las estructuras que asemejan raíces, consisten en rizoides no ramificados.
Los pteridofitos son cormofitos, en los cuales existe xilema y floema, que transportan tanto la savia bruta como elaborada en el órgano vegetativo (esporofito) que tiene también la función de reproducción vegetativa. No producen semillas, formándose los gametofitos a partir de esporas haploides independientes del esporofito. Comprenden las clases: Licopodios, Equisetos y Filicados o helechos.
Los equisetos, o colas de caballo, son plantas que proceden de antepasados muy difundidos durante el paleozoico que poseían portes arbóreos, y que se encontraban integrados en los bosques carboníferos. Desarrollan formas herbáceas, siendo en la actualidad el Equisetum el único género existente. Presentan tallos anuales que crecen a partir de un longevo, extenso y profundo rizoma. Externamente presenta una estructura con una serie de nudos y entrenudos muy evidentes; de los nudos salen hojas pequeñas y fotosintéticas, de los que a su vez surgen tallos que pueden ramificar. La epidermis es áspero y cortante al tacto debido a los depósitos de sílice que contiene. Algunas especies destacables son el Equisetum ramosissimum y Equisetum arvense (se le reconocen propiedades medicinales)
Por su parte, los licopodios son pequeñas plantas herbáceas; según la especie pueden ser acuáticas, anfibias o terrestres. Son igualmente descendientes de especies (lepidodendros) que cubrían los bosques carboníferos durante el paleozoico; algunas especies son verdaderas reliquias del Terciario. Habitan en climas tanto fríos como tropicales, incluso desérticos. Algunas especies muy extendidas son: Licopodium clavatum y Isoetes velata.
Los helechos son los pteridofitos más abundantes. Eran las denominadas antiguamente criptógamas vasculares, debido a su carencia de flores. Tuvieron su origen en el Devónico, siendo dominantes en los bosques del carbonífero. Son plantas perennes, ya que los rizomas desarrollan raíces adventicias de donde salen los frondes cada año. En zonas tropicales pueden desarrollar formas arbóreas de hasta 25 metros de altura, presentando aspecto de palmeras.
Los helechos presentan las hojas (frondes) erguidas, simples o compuestas. Habitualmente, las frondes están divididas o hendidas, y compuestas por foliolos independientes (pinnas), las cuales pueden estar a su vez divididas una o más veces (pinnulas), como ocurre en el helecho común. Típicamente habitan lugares frescos, húmedos y umbrosos. Los Leptosporangiados son la subclase más representativa de los helechos; éstos incluyen el helecho común (Pteridium aquilinum); helecho macho (Dryopteris filix-mas); helecho real (Osmunda regalis); polipodio común (Polypodium vulgare), etc. Determinados helechos, como el macho y común, se le reconocen propiedades medicinales.
Las gimnospermas son plantas espermatófitas (con semillas), que presentan los carpelos no diferenciados en ovario, estilo y estigma. Al contrario de las angiospermas, los óvulos y semillas no se forman en cavidades cerradas. Son plantas leñosas de porte variado, árboles o arbustos. Comprenden las clases Ginkgoineas, Coníferas, Cicadinas y gnetinas.
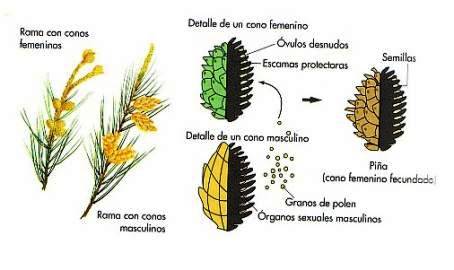
Son típicas plantas cormofitas, leñosas y muy ramificadas. Abundaban en el Jurásico y Cretácico formando tupidos bosques. Los órdenes Cordaitales y Coniferales comprenden a especies fósiles y actuales, que tuvieron su máximo desarrollo en el Mesozoico; algunos de ellos llegaban a los 30 metros de altura.
Muchas especies ya han desaparecido o reducido, pero se conservan otras como los pinos, de amplia distribución en la actualidad. En general se trata de árboles longevos, de gran porte, con hojas simples (trofofilos), numerosas, relativamente pequeñas y en su mayoría con forma acicular (cedros, pinos, enebros...) o escamosas (cipreses, tuyas o secuoyas) Salvo alguna excepción que presenta hojas caducifolias, éstas son normalmente perennes.
Aunque las características de las flores varían según los grupos, éstas carecen de periantio, siendo siempre unisexuales. Normalmente, las flores son monoicas, es decir, masculinas y femeninas situadas en la misma planta, aunque también se da el caso de flores dioicas (situadas en plantas independientes), como sucede por ejemplo en los enebros. En las formas más evolucionadas las flores femeninas (megasporangios) son muy reducidas, y están soldados por brácteas ovulíferas (escamas seminíferas) en los conos o estróbilos de las piñas. Según la especie, la semilla posee un ala para permitir la dispersión por el viento.
Por su parte, las flores masculinas (microsporangios) se agrupan en unos conos que incluyen numerosas hojas polínicas o microsporófilos, El polen contenido en los sacos polínicos poseen en ocasiones dos vesículas de aire, con objeto de facilitar la polinización anemógama (mediante el viento) Entre la polinización y la verdadera fecundación puede transcurrir más de un año. Las familias de las coníferas se dividen en: Pináceas, Cupresáceas, Taxodiáceas, Araucariáceas, Podocarpáceas, Taxáceas y Cefalotaxáceas.
En las gimnospermas, al igual que en las angiospermas, los gametofitos son microscópicos y crecen siempre a expensas del esporofito diploide. Sin embargo, a diferencia de aquellas, el órgano reproductor donde se encuentra el esporangio no es una flor con cáliz y corola, sino una estructura denominada cono.
Las semillas se desarrollan dentro del gametofito femenino, que es rico en nutrientes. Igualmente, se asemejan a las angiospermas en que faltan los flagelos en sus espermatozoides. Sin embargo, la característica evolutiva de las angiospermas de poseer flores hermafroditas no existe en las gimnospermas, salvo algunas excepciones, sino que los carpelos o los estambres siempre se hallan sobre flores distintas, y rodeadas o no por hojas estériles, que como sucede en el pino, forman la piña.
La mejor forma de exponer el proceso de reproducción de las gimnospermas es referirnos al pino, que aunque presenta variaciones secundarias en cuanto a la estructura floral y de reproducción, es en general representativo de estos vegetales.
Los pinos son especies monoicas. El ciclo reproductivo abarca dos estaciones vegetativas, como sucede en el resto de las gimnospermas. Existen flores masculinas portadoras de polen que son conocidas como cono de polen; están formados por unos múltiples microsporófilos que están dispuestos en hélice, con un pequeño pedículo y dos sacos polínicos, los microsporangios, donde se encuentran las células madre del polen. Estas células, por meiosis, forman gran cantidad de granos de polen (microgametofito), cuyo protoplasto se divide en dos células, una de las cuales es generativa y dará lugar a los espermatozoides y la célula tubular, encargada de formar el tubo polínico.
Los conos de semilla o flores femeninas están compuestos por escamas ovulíferas con dos óvulos cada una, consistentes en ramas reducidas y desplegadas en espiral rodeando un eje central; están situadas en la parte superior del árbol, es donde se produce la fecundación y la formación de la semilla. Durante la polinización el cono es pequeño, blando y verde. En cada una de sus escamas hay dos óvulos jóvenes, con la nucela o esporangio femenino, donde aparece una célula madre de la megaspora rodeada del integumento, presentando un orificio para la entrada del tubo polínico, al igual que en las angiospermas. En este estado y con las escamas del cono de semilla ligeramente separadas se produce la polinización. Es entonces cuando las escamas del cono se endurecen y se cierran en un cono compacto, que permanece así durante el verano hasta el invierno siguientes, que es cuando la célula madre de la megaspora se divide por meiosis formando cuatro megasporas; solo una de ellas se transformará en el gametofito femenino. Esta megaspora está formada por un tejido de almacenamiento y dos o tres arquegonios ovíferos, dentro del óvulo. En verano, al año aproximadamente de la polinización, finaliza este proceso y se produce la fecundación.
Los “frutos” y semillas de las gimnospermas son siempre anemócoros, es decir, se dispersan por el viento. La polinización se realiza por este medio, ya que los granos de polen poseen dos alas globosas. El polen es depositado en las proximidades del óvulo y queda fijado por una sustancia pegajosa que éste libera. Cuando sucede más tarde la germinación, la célula del tubo forma el tubo polínico, auque no se produce la división de la célula generativa hasta el verano siguiente, en que se produce la fecundación. El tubo polínico penetra a través del micrópilo y la célula generativa se transforma en una célula pedicular y otra somática, que originará los dos núcleos espermáticos. Uno de ellos es expulsado del tubo y degenera; el otro se une a la ovocélula formando un cigoto, que se desarrolla en la semilla, la cual una vez madura en otoño, sale del cono, cuyas escamas se separan y se encorvan.
Las semillas desarrollan un ala que les permite su dispersión por el viento. El embrión consta, como en las angiospermas, de un hipocotilo, varios cotiledones y un ápice caulinar. Se trata de un esporofito inmaduro que al germinar producirá una plántula. En las gimnospermas el ciclo dura dos estaciones vegetativas.
Los hongos son organismos que presentan características intermedias entre los animales y vegetales. Son unicelulares o pluricelulares con estructura de talo. Su nutrición es heterótrofa, es decir, no se nutren realizando la fotosíntesis (carecen de clorofila). Al igual que en los animales, su reserva energética es el glucógeno. Sus células, salvo en los grupos inferiores, no suelen estar desnudas, sino recubiertas por una membrana que suele ser de un polisacárido, la quitina.
Estas células se agrupan en talos filamentosos llamados hifas, cuyo conjunto constituye el micelio, que penetra en el sustrato. La reproducción puede ser sexual (por conjugación de hifas de sexos opuestos) o asexual (por medio de esporas) Al igual que ocurre con las plantas superiores, pueden existir individuos masculinos y femeninos, que morfológicamente pueden ser iguales o no; aunque se pueden dar hifas masculinas y femeninas dentro del mismo cuerpo vegetativo. También pueden multiplicarse por descomposición del micelio en células aisladas denominadas conidios.
La forma de vida de los hongos es básicamente de tres tipos:
- Saprofitos, que viven a expensas del material orgánico que descomponen retornándolo al suelo, por eso son más frecuentes en la hojarasca.
- Simbióticos, unidos a otro vegetal al que micorrizan ampliando la red capilar, lo que le aporta agua y nutrientes.
- Parásitos, que pueden llegar a matar árboles como en el caso de la grafiosis de los olmos.
Tienen, por tanto, una gran importancia ecológica, pues actúan como descomponedores junto a las bacterias o formando asociaciones simbióticas con otros vegetales, e incluso animales como los termes, proporcionándoles determinadas sustancias que no pueden producir por sí mismos. Tienen utilidad en la industria alimentaria y farmacéutica.
Los micofitos se clasifican en las clases Oomicetos, Ficomicetos, Ascomicetos, Basidiomicetos y líquenes (algunos autores incluyen los líquenes en una división independiente, al tratarse de la unión simbiótica de un alga y un hongo)
Los basidiomicetos son los hongos más evolucionados. A este grupo pertenecen las denominadas setas (orden Agaricales), así como los carbones de los cereales, royas, y otros que causan enfermedades en los vegetales.
La denominación basidiomicetos se refiere a que poseen unos basidios en el extremo de las hifas, donde se forman las esporas de origen sexual, siendo ésta la forma usual de reproducción, y cuyo ciclo es básicamente igual al de los ascomicetes; la reproducción asexual se da en basidiomecetos menos evolucionados. Se desarrollan de forma variable según se trate de hongos parásitos o saprófitos. Suelen ser saprófitos sobre materias orgánicas o suelos ricos en humus; y parásitos cuando se establecen sobre plantas superiores. Aunque pueden desarrollarse como parásitos de plantas acuáticas, existen muy pocas especies. Los órdenes más destacables de basidiomicetos son: Uredinales, Ustilaginales, Auriculariales y Agaricales.
Uredinales. Son las royas (llamadas así por el color rojizo de la lesión que producen) agrupan a parásitos de determinados vegetales, a los que pueden causar graves daños. La más conocida es la que se manifiesta en el trigo (Puccinia graminis)
Ustilaginales. Son las cenizas, tizones y carbones; abarcan a varios centenares de especies de hongos, huéspedes de las plantas superiores a los que suelen generar malformaciones. Atacan especialmente a las gramíneas. La infección se presenta en forma de unas masas de esporas con aspecto de hollín o polvo negro que terminan por asfixiar a la planta. Los más conocidos son los que atacan a los cereales, como el carbón del maíz (Ustilago maydis) o del trigo (Ustilago tritici)
Auriculariales. Son hongos gelatinosos, saprófitos de la madera, sobre la que se adhieren y concluyen pudriéndola. Destacan las tremelas (Tremella encephala y Tremella mesenterica) y la oreja de judas (Auricularia auricula Judae)
Agaricales. Se trata de las típicas setas; integra tanto a especies comestibles como venenosas. Son saprófitos sobre suelos o madera; en algunos casos son parásitos o constituyen micorrizas (asociación del hongo con las raíces de plantas superiores)
Especies comestibles son las del género denominado boleto (ejemplo del boleto edulis) y otras de alto valor gastronómico, como el el niscalo o rovellón (Lactarius deliciosus), la verdadera oronja o amanita de los césares (Amanita cesarea), la seta de cardo y chopo (Pleurotus eryngii y Pleurotus ostreatus), o el tan conocido champiñón (Psaliota campestris). Pero existen gran cantidad de especies que, si no son venenosas, no gozan de interés alguno en alimentación. Algunas especies son muy peligrosas por su toxicidad, ejemplo del boleto de Satanás (Boleto satanas); pero otras pueden incluso causar la muerte, como algunas pertenecientes al género Amanita: ejemplo de la vulgarmente llamada matamoscas (Amanita muscaria), la pantera (Amanita pantherina) o la oronja o cicuta verde (Amanita phalloides)
Las micorrizas
En las zonas mediterráneas de clima seco y continental es frecuente que los árboles y arbustos tengan sus raíces conectadas con el micelio de algunos hongos. Se trata de una relación de simbiosis en la que ambos salen beneficiados. Al desarrollarse las esporas, el hongo “infecta “ las raíces de la planta y éstas atrofian sus extremos formándose un engrosamiento de donde surgen finísimos filamentos de micelio que son capaces de introducirse por fisuras y poros de pocas micras. Así, la planta logra capturar agua y nutrientes de lugares donde no podría acceder sólo con sus raíces. El suministro de nitrógeno y de fósforo es también más intenso gracias al aprovechamiento del humus por el hongo. Los hongos reciben compuestos orgánicos del hospedante. Muchos de estos hongos sólo se observan al microscopio pero otros forman las setas (cuerpos de fructificación o carpóforos) que hacen fácil percatarse de su presencia.
En las micorrizas la asociación puede ser externa (ectótrofa) si se unen íntimamente en la zona superficial de la raíz, o interna (endótrofa), si las hifas de los hongos penetran en el interior de las células.
Un caso especial son las endomicorrizas de las orquídeas. Las diminutas semillas de estas plantas poseen muy poca cantidad de reservas y necesitan para germinar hongos simbióticos que les suministren agua, sales e incluso materia orgánica. También en las plantas adultas se encuentran hifas fúngicas en las células externas de la corteza de la raíz y, algunas especies, dependen siempre de los hongos porque son incapaces de realizar la fotosíntesis o apenas la realizan en estado adulto (Neottia, Corallorrhiza o Epipogium)
Las algas son de los primeros vegetales que poblaron la Tierra. Estos organismos
fotosintéticos llegaron a dominar el medio acuático pero, si querían expandirse,
tenían que conquistar el sustrato sólido. Al borde de las aguas, en los
apilamientos de algas arrojadas por las mareas, se daban las condiciones
apropiadas para que otros seres aprovechasen los nutrientes. Allí, debieron surgir los hongos, organismos capaces de
aprovechar la materia orgánica que no podían sintetizar como las algas por
carecer de clorofila o de pigmentos fotosintéticos.
los nutrientes. Allí, debieron surgir los hongos, organismos capaces de
aprovechar la materia orgánica que no podían sintetizar como las algas por
carecer de clorofila o de pigmentos fotosintéticos.
Como consecuencia de la coexistencia de esos dos tipos de organismos, aparecieron los líquenes, que son seres vivos formados por la unión de un alga y un hongo viviendo en simbiosis. Es decir, los dos organismos establecen una estrecha asociación en la que ambas especies se benefician. El hongo se encarga de proteger al alga de las radiaciones directas del sol y brindarle agua y sales minerales y, el alga, a su vez, realiza la fotosíntesis y proporciona al hongo alimento ya que la nutrición de éste es heterótrofa.
De este modo, mientras los mecanismos de vida de las algas y los hongos, por separado, tienen grandes limitaciones, la unión de ambos permite a los líquenes vivir en condiciones extremas y en lugares donde difícilmente podría desarrollarse una planta. Crecen en toda clase de superficies, como corteza de árboles, suelos y rocas, aunque son muy específicos del sustrato en que se desarrollan siendo incluso distintas las especies que viven en rocas silíceas de las que viven en rocas calcáreas. La falta de competencia y la ausencia casi total de depredadores, ha compensado su lento crecimiento, les ha permitido colonizar diversos territorios en el planeta y conseguir una amplia diversificación. Son organismos que soportan temperaturas entre los 70ºC y los –20ºC y que habitan en todo tipo de medio, desde desiertos a zonas antárticas. Se estima que existen alrededor de 13500 especies diferentes en el mundo.
Líquenes epífitos
Los líquenes pueden pasar rápidamente a un estado de vida latente cuando las circunstancias ambientales son desfavorables. Así, en momentos de desecación por el sol o el viento, suspenden su actividad y la recobran de nuevo con la lluvia o el rocío que absorben como si fueran papel secante ya que no poseen un órgano especializado para esa función.
En cuanto a su morfología, el talo o aparato vegetativo está formado por hifas entretejidas que rodean y protegen al alga. El talo puede ser homómero, si no se aprecia ninguna estratificación y las algas se hallan repartidas de manera uniforme entre las hifas, o heterómero, si se aprecian estratos: el córtex superior, formado por hifas y con función de protección, la capa gonodial próxima a la superficie, donde se encuentra el alga, la médula, formada por hifas y cavidades llenas de aire, y , a veces, el córtex inferior con función de protección y sujeción.
Las algas que componen los líquenes son de dos tipos: cianofitas o algas azules y clorofitas o algas verdes y la mayoría pertenecen a uno de estos tres géneros: Trebouxia, Trentepholia y Nostoc. Las cianofitas son capaces de fijar el nitrógeno atmosférico, algo que pocos grupos de seres vivos pueden hacer, por lo que los líquenes de los que forman parte tienen una gran ventaja. El hongo es, generalmente, un ascomiceto (hongos con reproducción sexual en los que las esporas se forman en el interior de ascos). Sólo el 2% restante corresponde a basidiomicetos y deuteromicetos.
Existen varios tipos de líquenes con aspecto muy distinto:
- los crustáceos (en forma de costra y fuertemente adheridos al sustrato por su cara inferior)
- los foliáceos (con forma de hojas o láminas y un borde que se separa bien del sustrato)
- los fruticulosos (con forma de arbustos y sujetos al sustrato por un único punto). Hay líquenes con talos compuestos que tienen una parte fruticulosa llamada podecio y una parte basal crustácea.
En la parte inferior del liquen pueden aparecer órganos de sujeción al sustrato como ricinas (“pelillos” muy frecuentes en los foliáceos) o discos de fijación (en algunos líquenes fruticulosos). También hay líquenes que presentan estructuras para facilitar el intercambio gaseoso como las pseudocifelas y las cifelas (interrupciones del córtex y capa algal para poner en contacto la médula con el ambiente externo)
Presentan los dos tipos de reproducción: sexual y asexual. La más común es por fragmentación del talo. Cuando el liquen se deseca, se vuelve frágil y el viento lo rompe y dispersa los fragmentos. Cada pedazo puede originar un nuevo talo en otro lugar. Muchos líquenes desarrollan propágulos vegetativos especializados, como los isidios y los soredios que, una vez desprendidos del talo, pueden originar otro nuevo cuando se dan las condiciones apropiadas. La reproducción sexual está a cargo del hongo que desarrolla ascocarpos en forma de apotecio o peritecio, con pequeños sacos llamados ascos donde se producen las esporas. Los apotecios, en forma de copa o disco abierto, se presentan sobre el talo, mientras que los peritecios , con forma globosa y un poro apical, se hunden en él.
En los ecosistemas, los líquenes son un importante componente, no sólo porque sirven de alimento y cobijo para multitud de seres vivos (en el caso del reno o el caribú, por ejemplo, constituyen más del 50% de su dieta invernal) sino porque posibilitan su desarrollo al favorecer la formación del suelo. Aunque en los bosques, el trabajo de estos seres primitivos pasa desapercibido ante el crecimiento de las plantas mas evolucionadas, en los lugares donde el hombre destroza el suelo, son los líquenes y los musgos los encargados de crear las condiciones para que se asienten las plantas vasculares.
La utilidad de los líquenes para el hombre es limitada. En algunos países nórdicos se consumen como alimento (Cetracia islandica) pero su uso más conocido es como fuente de colorantes. La primera tintura de tornasol usada para la determinación de pH fue obtenida a partir de líquenes. Recientemente se han descubierto propiedades antibióticas en algunas de las sustancias que producen. También suministran aceites esenciales que se utilizan en cosméticos y perfumerías.
Los líquenes son muy sensibles a la contaminación, especialmente frente al SO2,
y se utilizan como bioindicadores de ésta. La contaminación les afecta
drásticamente dañando sus células porque su estrategia de “esponja” hace que,
cada vez que se hidratan, todas las sustancias tóxicas que se han depositado
sobre ellos pasen a su interior. Además, a diferencia de las plantas vasculares,
no tienen cutículas, y absorben los gases en toda su superficie, resultando muy
sensibles a la contaminación gaseosa. Su gran longevidad y lento crecimiento
favorecen la capacidad de acumular diversos contaminantes, como los metales
pesados, en mayores concentraciones que en las plantas de su entorno. Así, la
ausencia de líquenes en las paredes o tejados de las ciudades es señal de
contaminación.
|
|
|
|